


O filo Echinodermata (do grego echinos, espinho + derma,
pele) inclui as conhecidas estrelas do mar, ouriços, bolachas da praia, pepinos
do mar, crinoide e ofiuroides. É composta por animais que apresentam simetria,
tipicamente pentâmera (cinco) ou mais radiais, derivadas secundariamente, e um
esqueleto calcário. Existem cerca de 6.000 espécies atuais descritas e 20.000
fósseis.
O endoesqueleto está localizado sob a
epiderme, os ossículos calcários dérmicos perfurados são
formados pelas células da mesoderme com espinhos ou espículas. Cada ossículo é
um único cristal e é separado por um tecido vivo. Este sistema é muito
diferente daquele característico dos esqueletos rígidos de outros animais, onde
os cristais encontram-se envolvidos por uma matriz orgânica. Ele permite um
aumento no tamanho sem a ocorrência de muda, pode ser rígido ou dobrado e
também pode ser servido por células vivas.
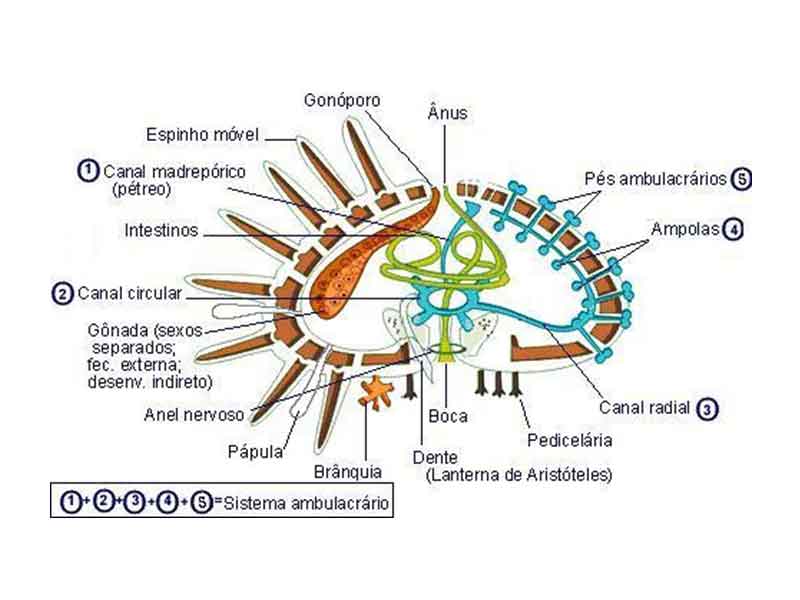
O sistema vascular aquífero do equinoderma
possui uma cavidade celômica central, que transporta água do mar pelo interior
do animal, e na qual estão inseridos os pés ambulacrais, extensões de paredes
finas da epiderme que são revestidas por mesoderme e conectadas com os canais
radiais. Na estrela-do-mar, por exemplo, a água marinha penetra através de uma
placa madrepórica, atravessa um "canal pétreo" central até chegar a
um canal anular e depois percorre canais radiais e penetra nos pés ambulacrais.
As funções dos pés ambulacrais são as seguintes:
a) Respiração através dos pés ambulacrais, brânquias
dérmicas, árvores respiratórias ou bursas dependendo da
classe do equinodema;
b) A coleta de alimentos; já o sistema digestivo na
maioria das vezes é completo, axial ou enrolado;
c) A locomoção também é realizada por esses pés e podem
servir como tentáculos sensoriais, através de receptores táteis.
O sistema nervoso é composto por um anel circum-oral e
nervos radiais; existem dois ou três sistemas de redes nervosas em determinados
locais do corpo. Isto sugere um grau de coordenação semelhante ao dos
cnidários, e ainda assim os grupos de células nervosas podem se transformar em
centros temporários de coordenação.

O tecido conjuntivo dos equinodermos apresenta
uma característica única e extremamente interessante. Ele é capaz de mudanças
rápidas e reversíveis na rigidez, frequentemente dentro de segundos, sob
controle nervoso. O mecanismo é uma alteração na viscosidade das fibras de
colágeno no tecido conjuntivo, causada por movimentos iônicos que modificam as
interações fracas entre as macromoléculas extracelulares. O processo é
controlado pela ligação do cálcio a células neurossecretoras. O resultado é que
o tônus muscular pode ser alterado sem gasto de energia envolvido na contração
muscular. Exemplos incluem o enrijecimento dos espinhos do ouriço do mar que
podem ancorar o animal, o enrijecimento e o relaxamento das paredes do corpo
das estrelas do mar e dos pepinos do mar, a autotomia dos braços dos ofiuroides
e a manutenção do tônus dos braços estendidos dos lírios do mar.
O Sistema circulatório (sistema hemal) muito
reduzido, com pouca ou nenhuma função na circulação e apresenta extensão do
celoma (sinus periemais).
Os equinodermos não são segmentados, o celoma não
é dividido ao longo do eixo corporal como em um anelídeo, mas ele existe como
um número de divisões separadas que persistem desde sua origem embrionária.
Além do sistema vascular aquífero principal, uma estrela-do-mar (por exemplo)
tem os seguintes celomas: um perivisceral grande, e a cavidade do sistema
hidrovascular contêm o celoma do tipo enterocélico (fluído celomático com
amebócito).

As larvas bilaterais livre-natantes nos animais deste
filo passam por vários estágios de desenvolvimento, sendo que alguns apresentam
desenvolvimento direto; a metamorfose leva ao estágio adulto ou a uma forma
subadulta: clivagem radial. As larvas se alimentam e se movem de uma
forma muito diferente dos adultos. A maioria possui larvas ciliadas, que
se alimentam por filtração e nadam no plâncton marinho.

Os gametas de equinodermas são estudados e manipulados em
laboratório, devido à facilidade de coleta e manipulação. A partenogênese
artificial foi descoberta primeiramente nos zigotos dos ouriços do mar.
Os equinodermas são quase que totalmente habitantes de
fundo e de substratos duros como pedras, rochas e corais, que também foram o
habitat de muitas formas extintas. Entretanto, algumas espécies de cada classe
invadiram fundos moles e adaptaram-se à vida na areia. Devido a sua estrutura
não são presas preferidas de outros animais, com exceção dos outros
equinodermos, alguns peixes e lontras marinhas.
Ainda não conhecem espécies de equinodermas parasitas,
mas alguns são comensais. O que foi visto é que uma variedade de animais vive
no interior destes animais, como as algas parasitas ou comensais, protozoários
cirripédios (cracas), copépodes, gastrópodes, bivalves, poliquetos, peixes e
outros equinodermas.
Os equinodermas constituem os deuterostomados (forma o
ânus e depois a boca), que são a segunda maior linha evolutiva do Reino Animal.
Ao contrário dos protostomados (primeiro forma a boca), a boca surge como uma
nova abertura localizada em oposição ao blastóporo, que forma o ânus. Os
ancestrais do filo eram simetricamente bilaterais e evoluíram, provavelmente
passando por um estágio séssil, que viviam no fundo do mar e possuíam um
esqueleto calcário massivo. A boca era virada para cima e o alimento era
coletado de todos os lados. A maioria dos equinodermos adultos atuais
move-se lentamente sobre o fundo do mar, coletando seu alimento com a boca,
localizada na superfície inferior dando origem as formas vágeis.
Classes:
Classe
Concentricycloidea

Possuem forma de disco e são menores de um centímetro de
diâmetro. São conhecidos como margaridas do mar, sendo descritas até o momento
apenas duas espécies. A simetria é pentarradial, mas não possuem
braços. Os pés ambulacrais estão localizados ao redor do disco. O sistema
hidrovascular é constituído de dois canais radiais concêntricos. O canal
externo é representado pelos canais radiais onde possuem os pés ambulacrais.
Apresentam um hidroporo que é homólogo ao madreporito, ligando o canal radial
interno com a aboral. Das duas espécies encontradas uma não possui trato
digestório, a superfície oral é coberta por um véu que é utilizado para
absorver nutrientes. Já a outra espécie possui estômago estreito em forma de
saco, mas o intestino e o ânus são ausentes.
Classe
Asteroidea

A classe Asteroidea (do grego aster = eidos, + forma =
ea, por) inclui os equinodermas que apresentam corpo com braços, pertencem às
estrelas do mar. Existem, tipicamente, cinco braços, mas em algumas estrelas do
mar podem existir vários. A maioria das estrelas do mar mede de 12 a 24 cm de
diâmetro e muitas apresentam cores intensas como vermelho, laranja ou azul.
A boca localiza-se no centro da superfície oral, cercada
por uma delicada membrana peristomal. A região ambulacral estende-se da boca
até a ponta de cada braço. O sulco ambulacral é rodeado por pés ambulacrais que
possuem espinhos móveis. Um grande nervo radial localiza-se entre as fileiras
dos ambulacrais. O nervo ocupa a extensão do celoma coberta por ossículos. Por
esse motivo que em asteroides e crinoides são chamados de aberto e nos demais
grupos são fechados.
A região aboral é áspera e espinhosa e em torno deste
possuem minúsculos apêndices em forma de mandíbulas, denominados pedicelários
cuja função é a de matar pequenos organismos que venham a fixar-se na
superfície do corpo. Tais organismos, assim como sedimentos, são varridos pelos
cílios do epitélio superficial. Pápulas (brânquias dérmicas ou brânquias
da pele) são projeções da cavidade celomática. E na mesma região aboral
encontram-se o ânus e o madreporito, circular. Um crivo calcário conectado ao
sistema hidrovascular.
A superfície dos asteroides é recoberta por um epitélio
ciliado. Abaixo da epiderme, o endoesqueleto apresenta placas calcárias
chamadas de ossículos calcários ligados uns aos outros por tecido
conjuntivo. Os ossículos são perfurados por uma rede de espaços geralmente
preenchidos por fibras e células dérmicas, esta estrutura é denominada deestereoma e
exclusiva de equinodermos.
As trocas gasosas e excreção principalmente a amônia é
realizada por difusão dos pés ambulacrais. O sistema vascular aquífero encontra
um sistema interno de canais derivados do celoma, abre-se para o exterior e,
poros (madreporito), que é aboral e perfurada por finos canais, os quais
convergem para um canal pétreo vertical que se estende oralmente até um canal
circular, embutido nos ossículos ao redor da boca.

Os canais radiais estendem-se para cada braço, passando
entre os ossículos de cima do sulco ambulacral. Cada canal lateral termina numa
ampola em direção aboral e num pé tubular em direção oral, que se projeta do
sulco ambulacral.
Muitas estrelas do mar estão adaptadas para viver em
fundos arenosos e podem até mesmo escavar. Estrelas de substratos moles possuem
ampolas duplas que exercem uma pressão suficiente para enfiar os pés tubulares
pontudos, sem ventosas, na areia.
O sistema nervoso pode ser considerado como
primitivo, estando intimamente associado à camada da epiderme. Um anel nervoso
está presente ao redor da boca e um nervo radial estende-se para cada braço.
Fibras do anel nervoso e dos nervos radiais fazem conexões com os neurônios do
plexo nervoso geral da epiderme.
Manchas oculares na extremidade de cada braço são
compostas por fotorreceptores e células pigmentadas e constituem o único órgão
sensorial. Células receptoras individuais estão presentes na epiderme corporal,
particularmente concentradas na epiderme dos pódios e nas margens dos sulcos
ambulacrais.
A troca gasosa e a excreção nas
estrelas do mar ocorrem através da superfície das pápulas e dos pódios e o
transporte interno processa-se através do líquido celomático. As pápulas das
estrelas que vivem em fundos macios estão protegidas da areia e sedimentos por
espinhos especiais em forma de tábuas, que fornecem um revestimento protetor
sob o qual alojam-se as pápulas e fluem as correntes ventilatórias. São estes
espinhos planos que explicam a aparência lisa de algumas estrelas do mar.
A boca abre-se num grande estômago cardíaco de paredes
espessas, que ocupa a maior parte do disco central. O estômago cardíaco abre-se
num pequeno estômago pilórico aboral. Um par de glândulas digestivas
localizadas em cada braço desemboca no estômago pilórico. Um curto intestino
estende-se do topo do estômago pilórico a um ânus não visível, localizado no
centro da superfície aboral. Associados ao intestino, existem cecos retais, que
são bolsas externas de função desconhecida. A maioria das estrelas do mar é
carnívora e necrófaga.
As estrelas do mar apresentam sexos separados e
nas duas gônadas desemboca em gonóporos ocultos na base dos braços. Os ovos são
lançados na água do mar, onde ocorre a fecundação. O desenvolvimento
processa-se no plâncton.
A larva é bipinária e depois é transformada em
braquiolária, vai para o fundo e se fixa ao mesmo, segue-se uma metamorfose
radical e complexa.
Classe
Crinoidea

A classe Crinoidea (do grego krinen, lírio + eidos,
forma) é representada pelos lírios do mar, possuem forma de uma flor presa a
uma haste. Geralmente possuem 70 cm de comprimento e a maioria habita os mares
relativamente profundos. Durante a metamorfose, os comatulídeos tornam-se
sésseis e pedunculados, mas com o passar do tempo passam a mover-se livremente.
A maioria vive em água profunda, mas os comatulídeos vivem em águas rasas,
especificamente no Indo Pacífico, Antilhas e
Caribe.
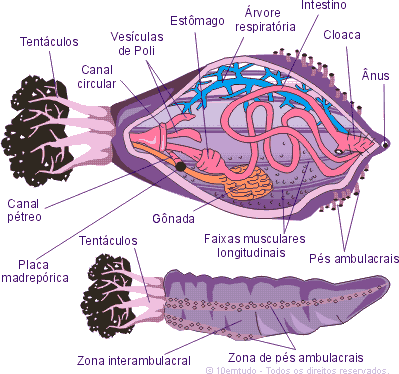
O disco do corpo ou cálice é coberto por uma pele chamada
de tégmen na qual há placas calcárias. A epiderme é pouco
desenvolvida e possui cinco braços flexíveis ramificados, formando outros
braços, cada um apresentará diversas pínulas laterais. O cálice e os braços
formam a coroa. As sésseis apresentam pedúnculos, que é composto por
placas e pode apresentar cirros.
Os sulcos de todos os braços convergem para a boca no
centro da superfície oral. A boca se abre no esôfago e este em um intestino com
divertículos que termina no ânus.
Os alimentos (pequenos organismos) são capturados pelos
sulcos ambulacrais (com auxílio dos pés ambulacrais) se abrem e levam o
alimento até a boca.
O sistema hidrovascular foi basicamente
descrito na aula passada na introdução dos equinodermos. O sistema nervoso
apresenta um anel oral e um nervo radial no interior de cada braço. Os órgãos
dos sentidos são escassos e primitivos.
Os sexos são separados, os gametas desenvolvem-se no
epitélio celomático dos braços e das pínulas e a desova é feita através da
ruptura da parede corporal. As larvas são denominadas bipinária e
livre-natantes durante algum período até a fixação e metamorfose.
Classe
Ophiuroidea

O nome da classe originou (do grego ophis, serpente +
eidos, forma), que inclui os ofiúros ou serpentes-do-mar. Pode-se citar como
exemplo, Amphipholis sp, Ophiothrix sp,Gorgonocephalus sp,
entre outras. O corpo dos ofiuróides, diferente dos asteróides, é composto
pelos braços e um disco central. Não possuem pedicelárias ou pápulas e seus
sulcos ambulacrais são fechados e cobertos por ossículos.
Os braços dos ofiuróides são preenchidos por um cordão de
grandes ossículos (vertebras). Não existe sulco ambulacral, mas os pódios
situam-se ao longo do lado oral dos braços. A articulação dos ossículos e a
musculatura dão aos braços uma grande mobilidade.
Os ofiuróides são especialmente comuns em habitats de rochas e corais, onde
vivem sob pedras ou em fendas e cavidades. Alguns, no entanto, vivem enterrados
em fundos arenosos. Os hábitos alimentares variam numa série de partículas,
seja pastejando ou filtrando. Os pódios dos ofiuróides não apresentam ventosas
nem ampolas. O estômago apresenta forma de um saco e o intestino é ausente.
Os ofiuroides não possuem pápulas; seus órgãos de
troca gasosa são bolsas, denominadas bursas, que estão dobradas para
dentro da parede corporal do disco oral, em ambos os lados da base de cada
braço. A corrente ventilatória é produzida por cílios ou por pressão de
mudanças no disco. O líquido celomático é o meio de transporte interno.
A maioria dos ofiuróides apresenta sexos separados e
as gônadas estão fixas ao lado celomático dos sacos bursais. Quando maduros, os
gametas são lançados nos sacos e liberados através das fendas. A larva é
denominada pluteus e a metamorfose ocorre antes dela ir para o fundo.
Não existe a fase da fixação, como nos asteroides. Muitas espécies incubam seus
ovos nos sacos bursais e os pequenos ofiuroides arrastam-se para fora por uma
das fendas.
Os sistemas hidrovascular, nervoso e hemal são iguais ao
da estrela-do-mar e cada braço contém pouco celoma.
Classe
Echinoidea

A classe Echinoidea (do grego echinos, ouriços do mar +
eidos, forma) inclui os ouriços do mar, cuja maioria está adaptada para a vida
em fundos duros, as bolachas da praia estão adaptadas para escavar na
areia. Em ambos os grupos, o corpo, que não apresenta braços, é esférico
ou discoidal. Os ossículos do esqueleto são placas achatadas, fusionados entre
si, formando um rígido esqueleto interno ou carapaça. A superfície do corpo é
recoberta por espinhos móveis encaixados sobre tubérculos da carapaça.
O corpo dos ouriços é esférico, com o polo oral, que
contém a boca, direcionada para baixo. Embora não existam braços, cinco áreas
ambulacrais, contendo pés tubulares radiais, vão do polo oral ao aboral. O ânus
abre-se no polo aboral, numa área circular, preenchida por pequenas placas,
denominada periprocto. Ao redor do periprocto, existem cinco grandes
placas genitais, cada uma contendo um gonóporo oculto. Uma das placas genitais
também funciona como madreporita.
Os longos espinhos móveis geralmente estão recobertos por
uma superfície de epitélio e articulados em tubérculos sobre a carapaça. Dois
arranjos musculares permitem que os espinhos fiquem rigidamente eretos ou
movimentem-se.
Os ouriços do mar usam os pódios, que podem estender-se
além dos espinhos, para a locomoção, do mesmo modo que os asteroides. Seu sistema
vascular aquífero também é muito semelhante. Entretanto, cada ampola do
lado interno da carapaça e o pódio correspondente do lado externo estão conectados
por perfurações duplas através dos ossículos. A locomoção também pode ser
auxiliada pelo movimento dos espinhos.
Alimentam-se de algas e animais incrustados, são raspados
ou mastigados por meio de movimentos complexos de um aparelho (conhecido como
lanterna de Aristóteles) que contém cinco dentes que se projetam através da
boca. O intestino é tubular e enrolado perto da carapaça.
Cinco pares de brânquias muito ramificadas estão
localizados a cada lado das áreas ambulacrais do polo oral, e são responsáveis
pela troca gasosa. O líquido celomático é bombeado para dentro e para fora das
brânquias. Como nos demais equinodermas, os pódios são finas extensões da
parede corporal, que contribuem, de alguma forma, para a troca gasosa.
Na superfície aboral, os pódios são muito largos e
achatados, modificados para troca gasosa. A densa cobertura de espinhos
estende-se além das brânquias podiais e impede que elas sejam obstruídas pela
areia. Cada uma dessas cinco áreas de pódios especializados denomina-se petaloide,
visto que se assemelham às pétalas de flores.
Nas bolachas da praia, finas partículas de matéria
orgânica, mas não grãos de areia, podem cair entre os espinhos. Essas
partículas, então, são levadas por cílios à superfície oral, onde passam por um
sistema ramificado de sulcos alimentares, que convergem para cinco outros
principais, correspondentes às áreas ambulacrais. Os sulcos alimentares são
contornados por pódios que levam massa alimentar coletada para a boca.
Classe
Holothuroidea

Os holoturoides ou pepinos do mar, são semelhantes aos
ouriços do mar por não possuírem braços. A maioria mede de 6 a 30 cm de
comprimento e possuem forma de um verme ou pepino. Habitam substratos duros,
vivendo sob ou entre, pedras ou em fendas nos corais. Esses pepinos
movimentam-se através de pés tubuIares.
A extremidade oral do corpo está circundada por
tentáculos, que representam pódios modificados ao redor da boca. Os tentáculos
estendem-se para fora e capturam plâncton ou material em suspensão na água do
mar e sedimento do fundo. Os que se alimentam de sedimento podem ser seletivos
ou não. Alguns não-seletivos que se alimentam de detritos deixam dejetos
visíveis sobre o fundo.
O aparelho digestivo dos pepinos do mar é
tubular e termina antes do ânus numa cloaca muscular, que tem participação na
troca gasosa. Essa troca é feita por estruturas tubulares ramificadas,
denominadas árvores respiratórias, que resultam de evaginações da parede da
cloaca e estendem-se pelo celoma. A ação de bombeamento da cloaca produz
uma corrente de água do mar, para dentro e para fora das árvores respiratórias.
As árvores respiratórias servem tanto para a respiração como para a excreção.
Alguns pepinos do mar possuem cachos de evaginações
tubulares na base da árvore respiratória. Esses túbulos, semelhantes a delgados
espaguetes, denominam-se túbulos de Cuvier e podem ser disparados,
exteriorizando-se do ânus. Eles são adesivos e alongam-se durante esse
processo. Um intruso ou predador que perturbe o pepino do mar é suficiente para
exteriorizar os túbulos que podem ficar emaranhados como uma armadilha mortal
de fios adesivos.
Os holoturoides possuem apenas uma gônada e o gonóporo
abre-se entre dois tentáculos. A larva bilateral é denominada auriculária,
possui faixas ciliadas locomotoras. A transformação em um jovem pepino do mar
ocorre antes deste ir para o fundo.
Artigo: Equinodermos do litoral do estado do Ceará
Artigo: Equinodermos do litoral do estado do Ceará
Gostei da matéria ajuda muito, gostaria de receber novidades sobre a materia.
ResponderExcluir